Tartalom
Az élővilágban a sejtes szerveződésnek két formája van, a prokarióta és az eukarióta szerveződés. A prokarióta sejtekben a maganyag és a citoplazma nincs elválasztva egymástól, míg az eukarióta sejtekben a maganyagot egy komplex nukleáris membrán választja el a citoplazmától. A sejtes evolúció során mindkét szerveződési forma sikeres volt, de amíg a prokarióta sejtek mindvégig megtartották egyszerű formájukat, az eukarióta sejtek bonyolult formákat hoztak létre. Az eukarióta sejtek evolúciójában fontos lépés volt a többsejtű szerveződési formák megjelenése, ami azután elvezetett a ma élő növények és állatok kialakulásához.
A többsejtű állatokban alapvetően két sejtpopuláció különíthető el, a szomatikus vagy testi sejtek, és a germinatív sejtek vagy ivarsejtek (gaméták). Az ivarsejtek az embrionális fejlődés korai szakaszában elkülönülnek az embrió testét felépítő szomatikus sejtektől. A szomatikus sejtek mitótikusan osztódnak, s az utódsejtek mindegyike a fajra jellemző diploid (2n) kromoszómakészlettel rendelkezik. Az ivarsejtek osztódása az ún. érési vagy meiótikus osztódás, melynek során az utódsejtek a fajra jellemző kromoszómakészletnek csak a felével rendelkeznek, vagyis haploidok (n).
A sejtosztódás (mitózis vagy meiózis) az eukarióta sejtek életének leglátványosabb szakasza. Kialakulnak a kromoszómák, melyek mozgása jól követhető az osztódások egyes szakaszaiban. Az osztódási fázisokat látszólag nyugalmi szakaszok követik, de ezekben a nyugalmi szakaszokban fontos molekuláris és funkcionális változások történnek a sejtekben. A sejt életében az osztódási és az ún. nyugalmi fázisok ciklikusan változnak. Az eukarióta sejtek ciklusa öt szakaszból, az M, G0, G1, S és G2 szakaszokból áll. Az egyes szakaszok hosszát, s így a sejt életfolyamatainak ciklikus változását az ún. ciklin-függő kinázok szabályozzák. Az egyes szöveteket felépítő sejtek életciklusa különböző hosszúságú. Általában azok a sejtek, amelyek a környezeti hatásoknak jobban kitett szövetek, pl. hámszövetek felépítésében vesznek részt rövidebb életciklussal rendelkeznek, ezért gyorsabban osztódnak. Azok a sejtek viszont, amelyek a szervezetben védettebb helyen vannak, mint pl. egyes kötőszöveti sejtek vagy az izomsejtek, hosszú életciklussal rendelkeznek, vagyis ritkábban osztódnak. Extrém esetben, amikor a szövetek, szervek egyensúlyi működésének alapfeltétele a sejtek állandósága, mint pl. az idegsejtek esetében, a sejtek egyáltalán nem osztódnak, akár egész életüket a ciklus G0 fázisában töltik. Különleges jelek, pl. növekedési faktorok hatására azonban ezek a sejtek is kimozdíthatók ebből az állapotból, s rábírhatók az osztódásra.
A szöveteket alkotó sejtek sejtciklus hosszától függően szoktunk beszélni ún. statikus sejtpopulációkról, ahol a sejtek már nem osztódnak. Ilyenek pl. az idegsejtek vagy a vázizomszövet sejtjei. Statikus sejtpopulációknak tekintjük a ritkán osztódó sejteket is, mint pl. a szívizomsejteket vagy a simaizomsejteket.
Stabil sejtpopulációkat alkotnak azok a sejtek, amelyeknek a mitótikus aktivitása alacsony, de bizonyos körülmények, pl. sérülések, regeneráció során ez felgyorsulhat. Ide tartoznak pl. a kötőszövetek rostképző sejtjei, mint pl. a fibroblasztok, oszteoblasztok és a különböző szerveket borító hámsejtek. A megújuló sejtpopulációk magas mitótikus aktivitással rendelkező sejteket tartalmaznak. Ilyenek pl. a vérsejtek, a többrétegű hámok alsó rétege vagy a bélcsatorna lumenét borító hám.
Az eukarióta sejtek két fő alkotórésze a citoplazma és a sejtmag. A citoplazmaorganellumok legnagyobb része membránnal körülvett struktúra, de vannak ún. nem-membrán citoplazmaorganellumok is. A membránorganellumok közé soroljuk a plazmamembránt, a durva felszínű (DER) és a sima felszínű (SER) endoplazmatikus retikulumot, a Golgi apparátust, a mitokondriumot, a lizoszomát, endoszomát és a peroxiszomát. A nem membránorganellumok közé soroljuk a mikrotubulusokat, mikrofilamentumokat, átmeneti vagy intermedier filamentumokat, a citoszkeletális organellumokat mint pl. a centriólumot és a riboszómákat.
A plazmamembrán elsődleges feladata, hogy elkülönítse a sejt belső környezetét a külső környezettől. Emellett még számos sejtélettani funkcióban játszik meghatározó szerepet. A plazmamembrán teljes vastagsága általában 8-10 nm. Az állati sejtek legnagyobb részénél a plazmamembrán külső felszínén egy szénhidrát réteg, az ún. glikokalix található. A glikokalix cukormolekulái kovalens kötéssel kapcsolódnak a plazmamembrán fehérjéihez és lipidjeihez. A plazmamembrán szerveződését az ún. félfolyékony mozaik modell segítségével érthetjük meg. A plazmamembrán, s a biológiai membránok általában foszfolipidekből, koleszterinből és fehérjemolekulákból épülnek fel. A membránt felépítő lipidmolekulák két rétegben helyezkednek el úgy, hogy a zsírsavláncuk egymás felé néz. Ennek következtében a membrán belső része hidrofób, vagyis víztaszító. A plazmamembrán citoplazmatikus és extracelluláris felszínét a lipidmolekulák poláris, feji vége képezi. A membránfehérjék egy része, az ún. integráns membránfehérjék hidrofób részeikkel beágyazódnak a lipid kettősrétegbe, vagy sok esetben át is érik azt. A plazmamembrán extracelluláris felszínén a fehérjékhez és a lipidekhez szénhidrát molekulák kapcsolódnak, így azok glikoproteineket, illetve glikolipideket alkotnak.
A membránproteineket funkciójuk alapján hat csoportba szokták sorolni. A pumpák az ionok aktív transzportját irányítják. A csatornafehérjék elektromos vagy koncentrációgrádiens mentén biztosítják bizonyos ionok mozgását. A receptor proteinek specifikus kötőhelyeket biztosítanak az extracelluláris térben lévő anyagok (ligandok) számára. Receptorproteinek közvetítik pl. a különböző hormonok vagy antitestek hatását. A transzducer proteinek részt vesznek a receptor-ligand kötődésben, illetve a másodlagos hírvivő molekulák aktiválásában. Vannak membránhoz kötött enzimek, mint pl. egyes ATP-ázok. Ezen kívül a különböző struktúrfehérjék a sejteknek egy viszonylag állandó szerkezetet biztosítanak.
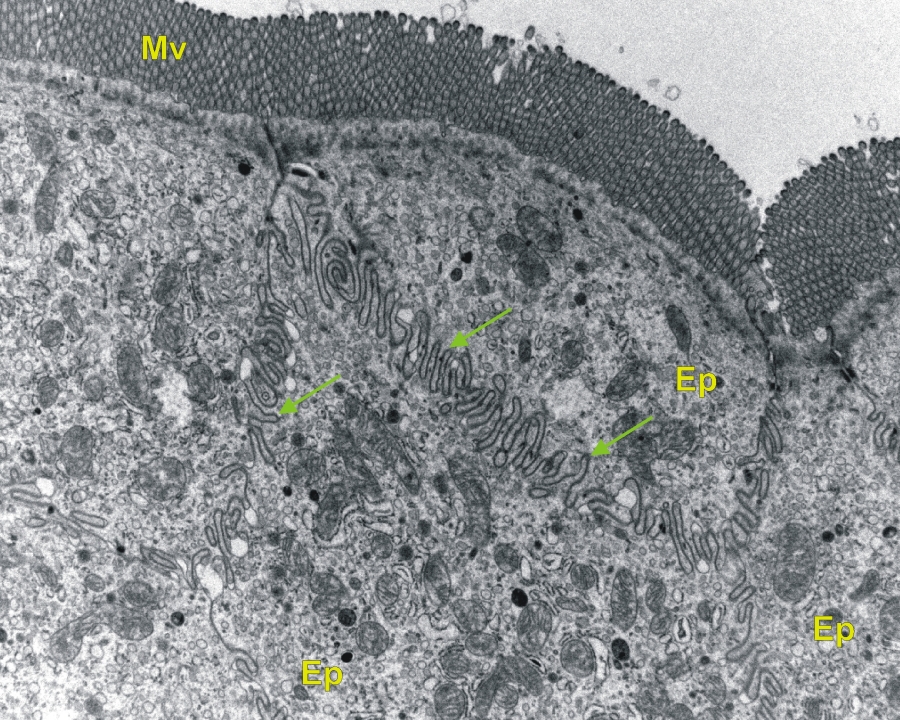
2/1. ábra Patkány vékonybeléről készült transzmissziós elektronmikroszkópos felvétel . A felvételen a bélfal epitheliumát (lamina epithelialis) alkotó hámsejtek láthatók, az apikális felszínükön mikrovillusokkal (nyilak-sejtmembrán interdigitáció, Mv-mikrovillus) 4600x
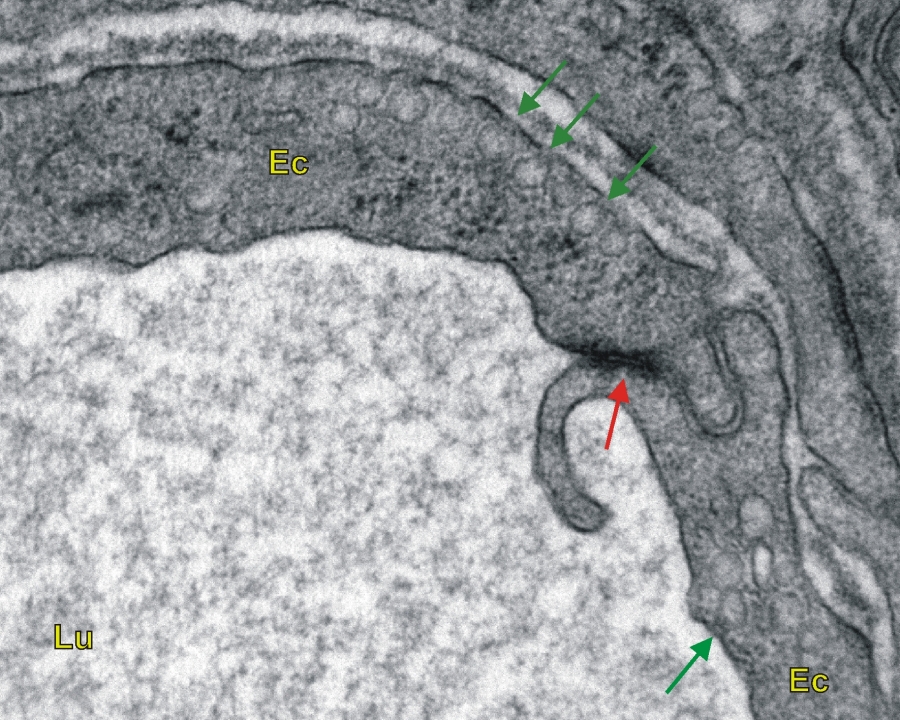
2/2. ábra Patkány vékonybeléről készült transzmissziós elektronmikroszkópos felvétel. A felvételen egy kapilláris endotélsejtjei láthatók transzportvezikulákkal. (Ec-endotelsejt, Lu-kapilláris lumene, piros nyíl-sejtkapcsolat, zöld nyilak-transzportvezikulák) 46000x
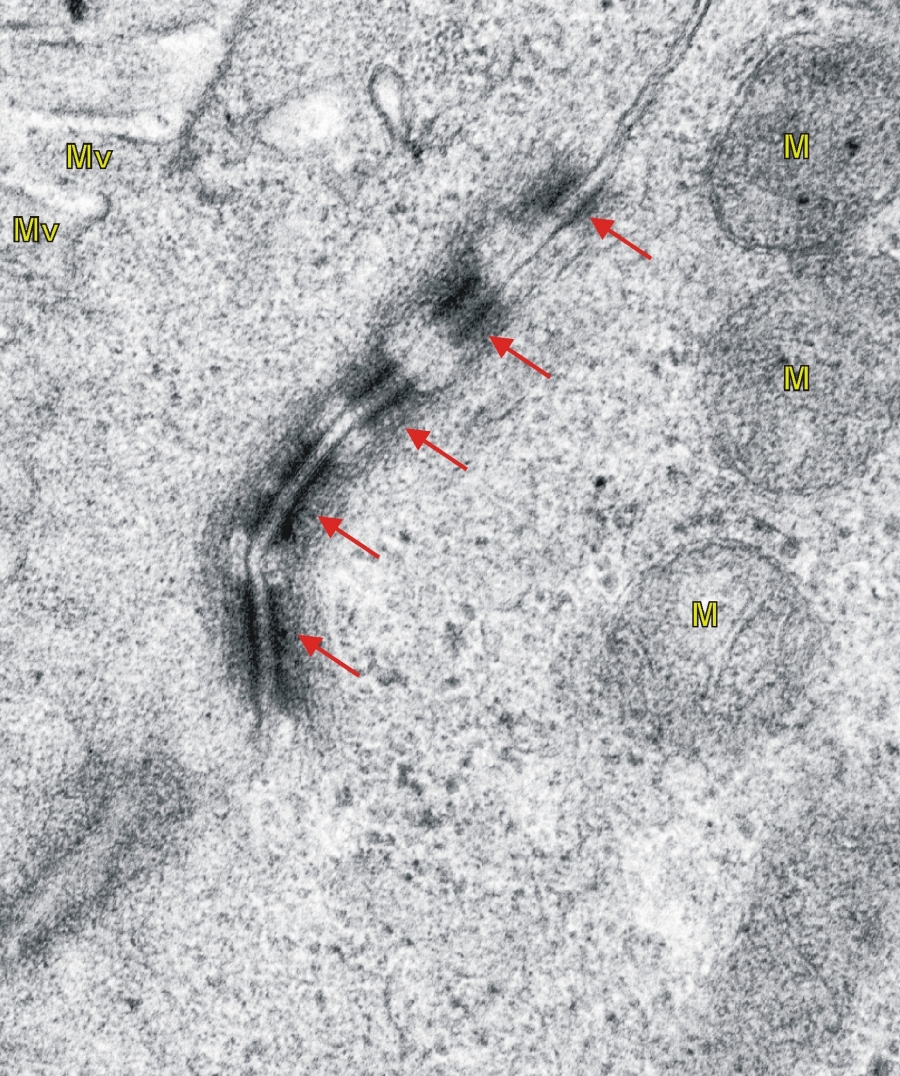
2/3. ábra Bélhámsejtek közötti adherens kapcsolatok (piros nyíl) transzmissziós elektronmikroszkópos képe. (M-mitokondrium, Mv-mikrovillus) 46000x