Tartalom
Az ektodermális eredetű idegszövetet idegsejtek és gliasejtek komplex hálózata építi fel. Emellett az idegszövetben sok véredény is található. Az idegszövet kapillárisaiban az endothelsejtek között szoros sejtkapcsolatok (zonula occludens) alakulnak ki, amelyek megakadályozzák, hogy a vérben keringő anyagok szabadon kijussanak az extracelluláris térbe. Így az idegszövet kapillárisainak endotheliuma, a nem ablakos vagy nem fenesztrált endothelium meghatározó morfológiai eleme a központi idegrendszerben a vér-agy-, illetve a perifériás idegrendszerben a vér-ganglion gátnak. A neuronok rendkívül magas metabolikus aktivitással rendelkeznek. Ez egyrészt magyarázható azzal, hogy a sejteknek a szerteágazó nyúlványrendszer miatt igen nagy a felszíne, másrészt a neuronok folyamatos működéséhez szükséges elektrokémiai grádiens fenntartásához is sok energiára van szükség. Ez a magas metabolikus aktivitás jól tükröződik a neuronok ultrastruktúrájában.
A neuron általános szerveződése
A sejttest vagy perikaryon tartalmazza a sejtmagot és a sejt fenntartásához szükséges sejtorganellumok nagy részét. A sejttestből egy hosszú nyúlvány, az axon lép ki, amely más sejtek számára közvetíti a jeleket. A sejttestből számos rövidebb nyúlvány is kilép, ezek a dendritek. A dendritek jelentősen megnövelik azt a felszínt, amelyen keresztül az adott sejt más sejtekkel kommunikálni képes. A sejtek között speciális kapcsolatok, szinapszisok alakulnak ki. A szinapszisok teszik lehetővé a különböző neuronok közötti közvetlen kommunikációt.
Neurocitológia
A nukleusz nagy, általában kerek, nagy központi elhelyezkedésű nukleolusszal. A kromatin állomány főleg eukromatint tartalmaz, ami a magas transzkripciós aktivitás morfológiai jele. A neuronokban nagyon kiterjedt DER található, ami az aktív fehérjeszintézisre utal. Ez a nagymennyiségű DER hematoxilin-eozin festett fénymikroszkópos preparátumokon lila szemcsékként látszik a citoplazmában. Ezeket a szemcséket leírójuk után Nissl anyagnak nevezzük. Sok Nissl anyag található a perikaryonban és a dendritekben, de az axonban nincs. A neuronban jól fejlett Golgi apparátust is találunk, ami a fokozott szekréciós aktivitásra utal, míg a nagy mennyiségű mitokondrium a sejt nagy energiaigényét elégíti ki.
Mivel a neuronokban az intenzív szekretorikus funkció miatt nagy a membránkörforgás, citoplazmájukban igen sok lizoszóma is található. Lipofuszcin tartalmú reziduális testek is gyakran előfordulnak, különösen az öregedő sejtekben. A neuronok citoszkeletonja különlegesen jól szervezett, hiszen ennek a feladata fenntartani a neuronok jellegzetes morfológiáját, s ezen belül a különösen hosszú, akár 1 m hosszúságú axonokét is. A sejtalak fenntartása mellett a mikrotubuláris rendszer meghatározó szerepet játszik az axonterminálisok felé (anterográd), illetve az axonterminálisokból a perikaryon felé (retrográd) tartó vezikuláris transzportban. A mikrotubulusok mellett jellegzetes intermedier filamentumok a neurofilamentumok, illetve az aktin mikrofilamentumok, s a hozzájuk kapcsolódó fehérjék alakítják ki a citoszkeletális hálózatok. Az aktin filamentumok amellett, hogy fontos szerepet játszanak a sejt alakjának a fenntartásában, igen jelentősek az idegsejtek növekedésében, migrációjában, az axonok növekedésében is.
A neuronok morfológiája
A neuronok változatos alakúak, s alakjuk jól tükrözi funkciójukat. A motorneuronok nagy sejttestje biztosítja a hosszú axon számára szükséges energiát. A motorneuronoknak sok dendritjük is van, így ezek a neuronok multipoláris neuronok. Az érző neuronok általában egynyúlványú, ún. unipoláris neuronok. A gerinces állatok elsődleges érző neuronjaiban ez az egy axon a sejttestet elhagyva kettéválik (pszeudounipoláris neuron). Hosszabbik, ún. perifériás nyúlványa az érzőreceptorokhoz megy, míg rövidebb, ún. centrális nyúlványa belép a központi idegrendszerbe. Az interneuronok általában kicsi, de jellegzetes morfológiájú (csillagsejtek, kosársejtek, stb.) neuronok, amelyek a központi idegrendszerben és a bélidegrendszerben helyi kapcsolatokat alakítanak ki. Az interneuronok nagy része bipoláris neuron, egyik nyúlványa axon, a másik dendrit. Az alaptípusok mellett nagyon sok, egy-egy idegrendszeri területre jellemző, jellegzetes morfológiájú neuron van. Ilyen pl. a kisagykéreg Purkinje-sejtje vagy a nagyagykéreg piramissejtje.
Axonális transzport
Ahhoz, hogy a hosszú axon metabolizmusa egyensúlyban legyen, a különböző sejtorganellumokat, enzimeket és metabolitokat folyamatosan szállítani kell a sejttestből a terminálisok felé és vissza. Az enzimek és a citoszkeletális fehérjék az ún. lassú axonális transzporttal szállítódnak az axonban. Ennek a mechanizmusa valójában nem ismert, sebessége napi 1-5 mm. A különböző membránorganellumok, mint pl. a transzmitter vezikulák napi 400 mm-es sebességgel, a már előbb említett anterográd gyors axonális transzporttal szállítódnak. Ezt a folyamatot a mikrotubulusok irányítják, a kinezin nevű motorprotein segítségével. A különböző membránokorganellumok visszaútja a sejttestbe, a retrográd gyors axonális transzporttal 300 mm/nap sebességgel történik. Ezt a folyamatot is mikrotubulusok irányítják, de a retrográd transzportban a dinein molekula játszik fontos szerepet.
Szinapszis
A szinapszis egy speciális sejtkapcsolat, ami a sejtek között közvetlen kommunikációt biztosít. A szinapszisoknak két alapformája létezik, nevezetesen a kémiai szinapszis, amikor az egyik sejt egy morfológiailag jól körülhatárolt területen ingerületátvivő, neurotranszmitter anyagot szekretál, s ezt az anyagot egy másik sejt nagyon specifikus módon képes felvenni. A másik alapforma az elektromos szinapszis. Az elektromos szinapszis morfológiai alapja a réskapcsolat (gap junction).
A kémiai szinapszis szerkezete
Az axon terminális része kiszélesedik és kialakítja a preszinaptikus felszínt. Ez egy kb. 20 nm szélességű szinaptikus résen keresztül kapcsolódik a célsejthez, vagyis a posztszinaptikus sejthez. Mind a pre-, mind a posztszinaptikus membrán speciális membránfehérjéket és receptorokat tartalmaz, amelyek fontos szerepet töltenek be a neurotranszmisszió folyamatában. Ultrastrukturálisan mindkét membrán egy kicsit megvastagodott. A preszinaptikus rész mitokondriumokat, mikrotubulusokat, neurofilamentumokat és 40-65 nm átmérőjű, változatos alakú és denzitású membránhoz kötött vezikulákat tartalmaz. Immunohisztokémiai módszerrel azonosítható a vezikula tartalma. A vezikulák a transzmittermolekulákon kívül más specifikus fehérjéket is tartalmaznak, mint pl. synaptophysin vagy chromograninek, amelyek az ingerületátvivő molekulák végső kialakításában, becsomagolásában játszanak fontos szerepet. A szinapszisok különböző kombinációban alakulnak ki. Az axonok és dendritek közötti kapcsolat az axo-dendritikus, az axonok és a sejttest közötti az axo-szomatikus, míg két axon közötti, az axo-axonikus. Egy célsejten különböző axonok végződhetnek. A célsejt az axonokon beérkező információkat integrálja, s kialakítja a megfelelő választ. A motoros axonvégződés és a vázizomrost membránja között kialakuló speciális szinapszis, a motoros véglemez.
Gliasejtek
A perifériás idegrendszerben a legáltalánosabban elterjedt gliasejttípus a Schwann-sejt, ami a perifériás idegek táplálásában, valamint a környezettől való elszigetelésében, mielinizációjában játszik fontos szerepet. A perifériás idegdúcok gliasejtjei a szatellita sejtek. A bélidegrendszer ganglionjaiban lévő enterogliasejtek felépítésükben és működésükben nagyon hasonlítanak a központi idegrendszer asztrogliasejtjeire.
A központi idegrendszer gliasejtjei lehetnek mikroglia és makroglia sejtek. A makroglia közé soroljuk a mielinizációban fontos szerepet játszó oligodendrocitákat, valamint a különböző típusú plazmás és rostos asztrocitákat. Ez utóbbiaknak az idegsejtek táplálásában és a vér-agy gát kialakításában van meghatározó szerepük. A Hortega-féle mikroglia vagy mesoglia, amely az idegszövet egyetlen mezodermális eredetű sejtje, a mononukleáris fagocitáló rendszer tagjaként, az idegrendszer immunológiai védelmében játszik fontos szerepet.
A mielinizáció
Kábelszerű vezetés esetén a vezetés sebessége függ a kábel vastagságától. Így az óriásaxonokon nagyobb sebességgel terjed az akciós potenciál. Az óriásaxonok fenntartása ugyanakkor rendkívül energiaigényes. Ezért az idegrendszer evolúciója során a kábelszerű vezetés mellett kialakult az ún. szaltatorikus vezetés, ami a mielinizációnak köszönhető. A mielinizációt a központi idegrendszerben az oligodendrocita sejtek, míg a periférián a Schwann-sejtek végzik. Az oligodendrociták több, egymás közelében lévő axont is mielinizálnak, míg a Schwann-sejtek csak egy axont. Az oligodendrocita sejtek nyúlványaikkal körbefonják az axonokat. A periférián viszont úgy kezdődik a mielinizáció, hogy az egyetlen axont körbefonja a Schwann-sejt külső membránja, így kialakul a belső mesaxon. Ezután a sejt többször körbecsavarodik az axon körül. A feltekeredő membránok közül a citoplazma kiszorul, a membránok fúzionálnak és kialakítják a külső mesaxont. A Schwann-sejtek membránja speciális lipidekben, pl. galaktocerebrozidokban gazdag. A mielinhüvely nem folyamatos az axon teljes hosszában, hanem 1-2 mm-es egységekből tevődik össze. Minden egység egy-egy Schwann-sejtet jelent. A kis egységek közötti tér a Ranvier-féle befűződés. Itt találhatók a feszültségfüggő ioncsatornák, így ezeknek meghatározó szerepük van az ingerület terjedésében. Az ingerület befűződésről befűződésre ugrásszerűen, szaltatorikusan terjed tovább. Vannak olyan idegrostok, amelyek nem trartalmaznak velőshüvelyt, ezeket velőtlen rostoknak nevezzük. Túlnyomó többségük azonban különböző módon kapcsolatba lép a környezetükben lévő Schwann-sejtekkel. Az idegrostok az idegszövetben hálózatot, neuropilemát vagy kötegeket képeznek. A kötegeket a központi idegrendszerben pályának, a periférián idegeknek nevezzük. A gliasejteknek a mielinhüvelyképzés mellett fontos feladata van az idegsejtek speciális anyagcseréjének az ellátásában. Ezen túlmenően egyes fagocitózisra képes sejtek képesek immunsejtekként is viselkedni. Speciális gliasejteket, pituicitákat találunk a hipofízis hátsó lebenyében, Müller-féle sejteket a retinában, és ependimasejteket az agykamrákban.
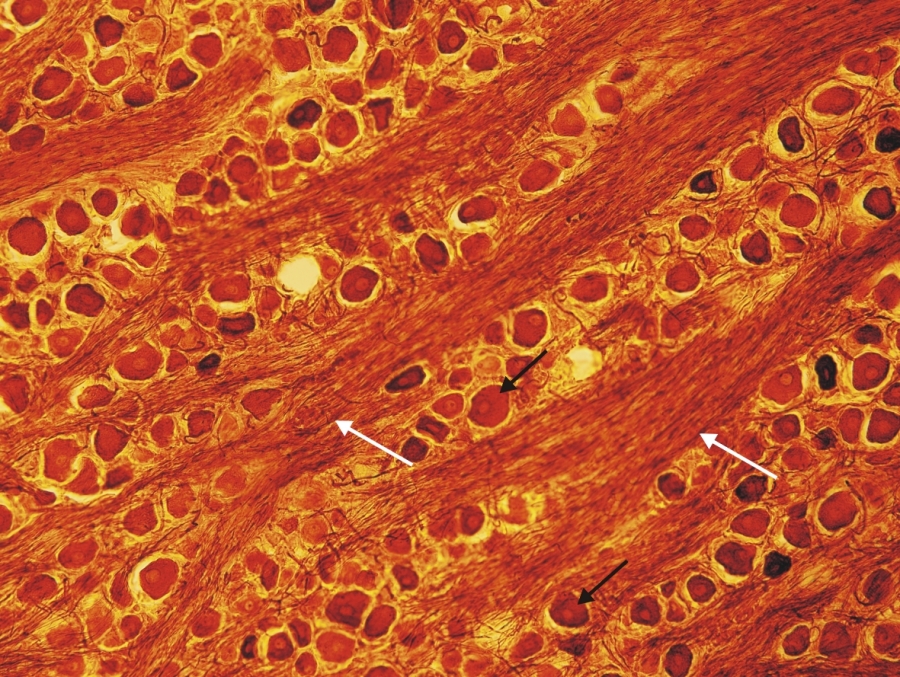
Fénymikroszkópos felvétel kutya Gasser-dúcából készült metszetről. (fehér nyíl-idegrostok, fekete nyíl-pszeudounipoláris neuron) 200x
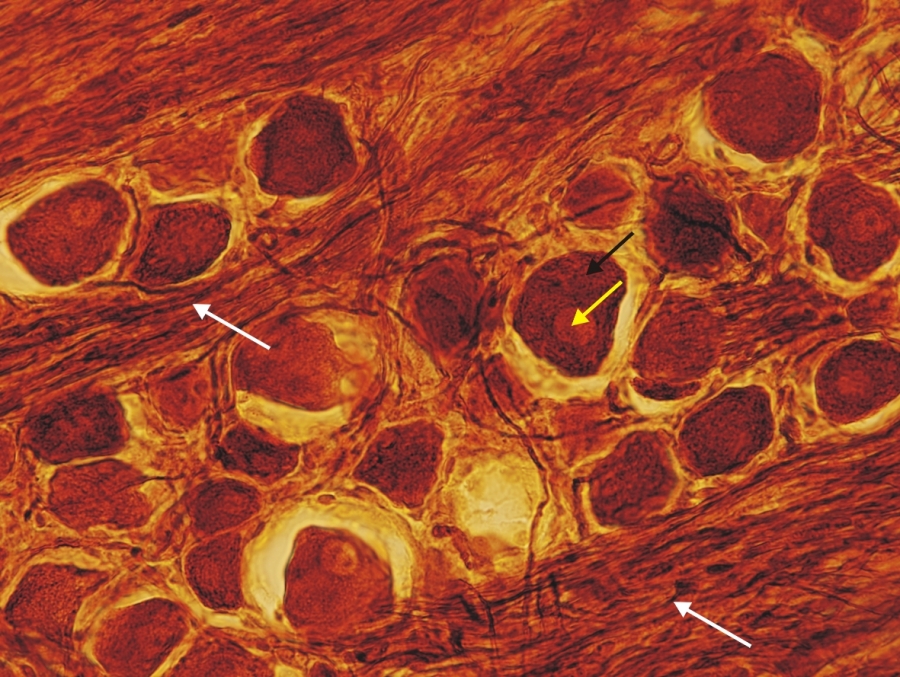
Fénymikroszkópos felvétel kutya Gasser-dúcából készült metszetről. ( fehér nyíl-idegrostok, fekete nyíl-pseudounipoláris neuron szómája, sárga nyíl-pszeudounipoláris neuron sejtmagja) 630x
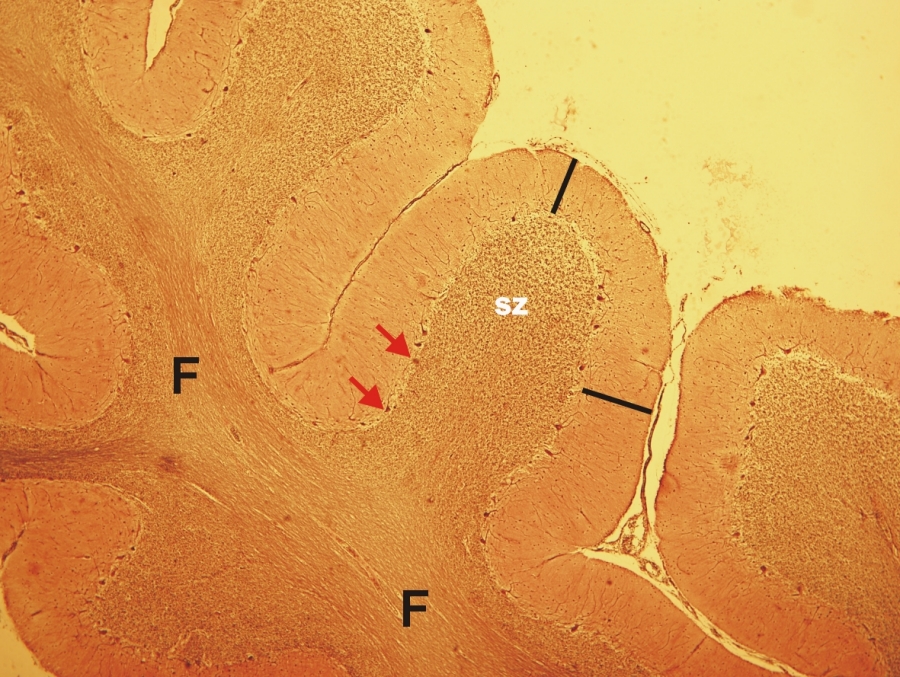
Fénymikroszkópos felvétel humán kisagyból készült metszetről. (F-fehérállomány, sz-szemcsesejtek rétege, piros nyíl-Purkinje-sejtek rétege, fekete vonal-molekuláris réteg) 50x
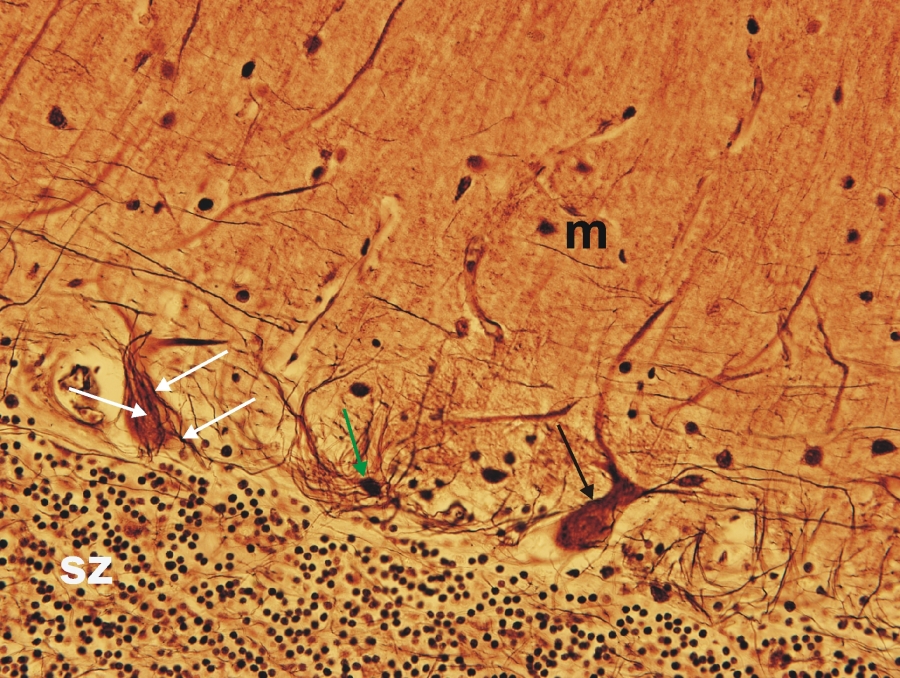
Fénymikroszkópos felvétel humán kisagyból készült metszetről. (sz-szemcsesejtek rétege, fekete nyíl-Purkinje-sejt, zöld nyíl-kosársejt, fehér nyíl-kosársejtek nyúlványai, m-molekuláris réteg). 400x
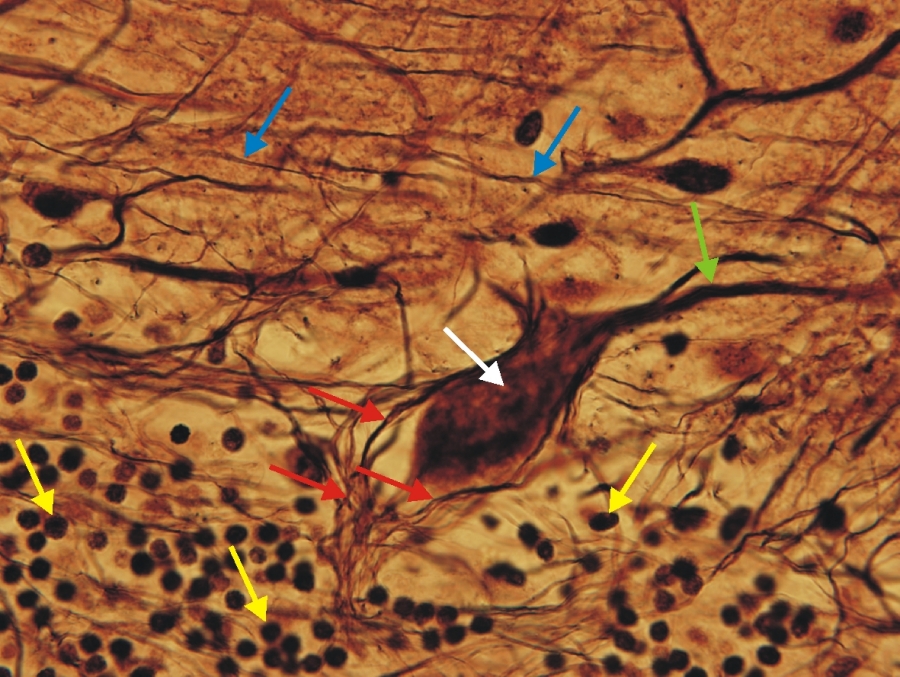
Fénymikroszkópos felvétel humán kisagyból készült metszetről. (sárga nyíl-szemcsesejtek, fehér nyíl-Purkinje-sejt sejttestje, zöld nyíl-Purkinje-sejt dendritje, piros nyilak-kosársejtek nyálványok, kék nyilak-parallel rostok) 1000x

Fénymikroszkópos felvétel humán cerebrumból készült metszetről. (nyíl-piramissejt) 100x

Fénymikroszkópos felvétel humán cerebrumrumból készült metszetről. (fehér nyíl-piramissejt sejttestje, sárga nyíl-sejtmag, zöld nyíl-apikális dendrit, zöld nyílhegy-bazális dendrit ) 400x

Fénymikroszkópos felvétel nyúltvelőből készült metszetről. (fehér kör-magcsoport, fehér nyilak-rostok)
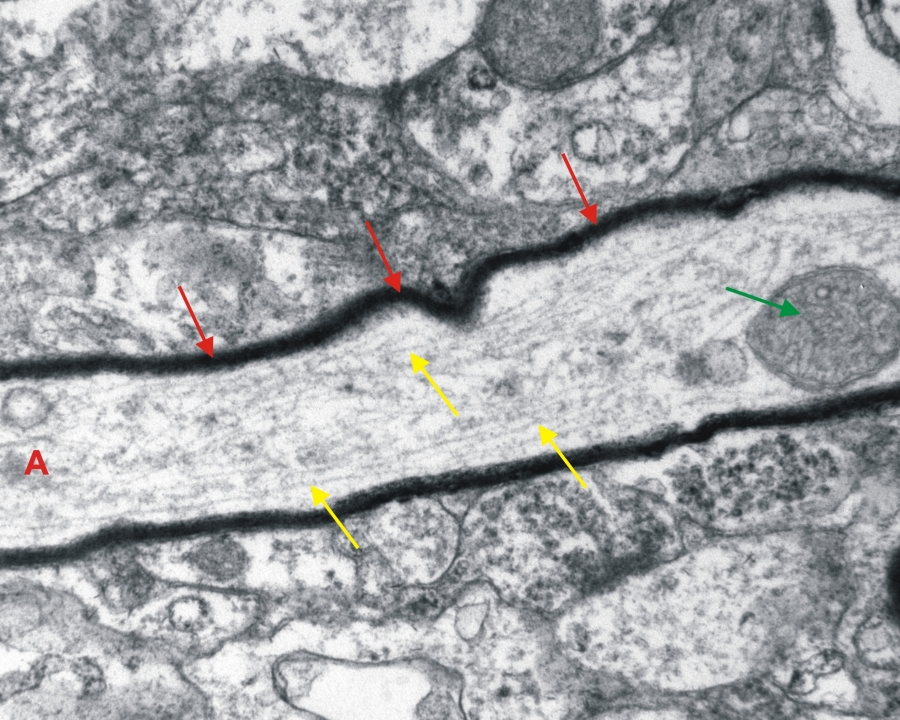
Transzmissziós elektronmikoszkópos felvétel enterikus ganglionról patkány vékonybélben. (A-axon hosszmetszete, piros nyíl-mielinhüvely, zöld nyíl-mitokondrium, sárga nyíl-citoszkeletális filamentum) 25000x
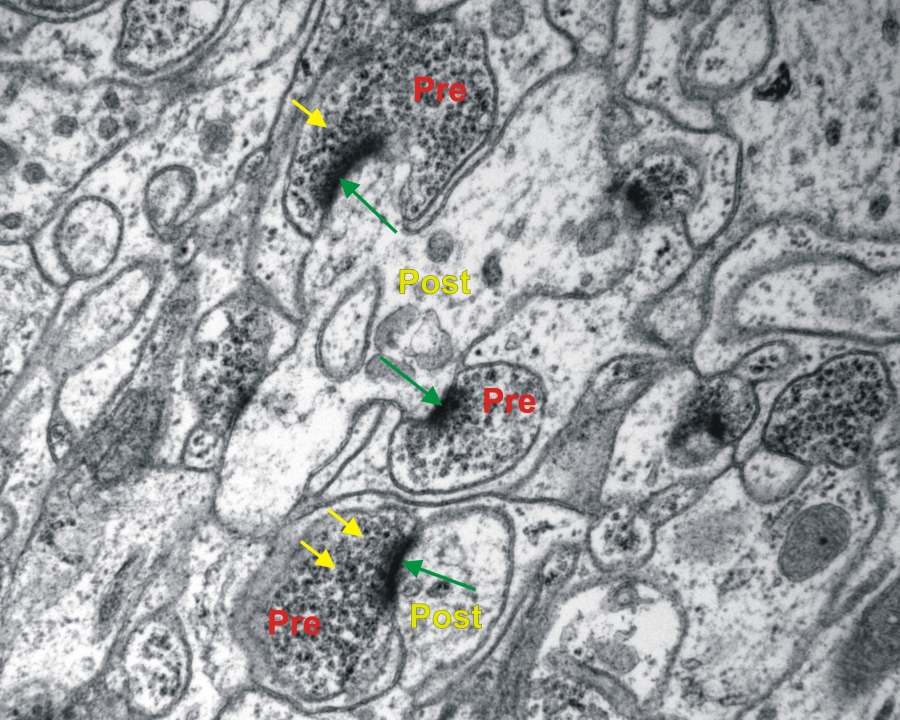
Transzmissziós elektronmikoszkópos felvétel entericus ganglionról patkány vékonybélben. (zöld nyíl-kémiai szinapszis, Pre-preszinaptikus sejt, Post.posztszinaptikus sejt, sárga nyíl-szinaptikus vezikulák) 19000x
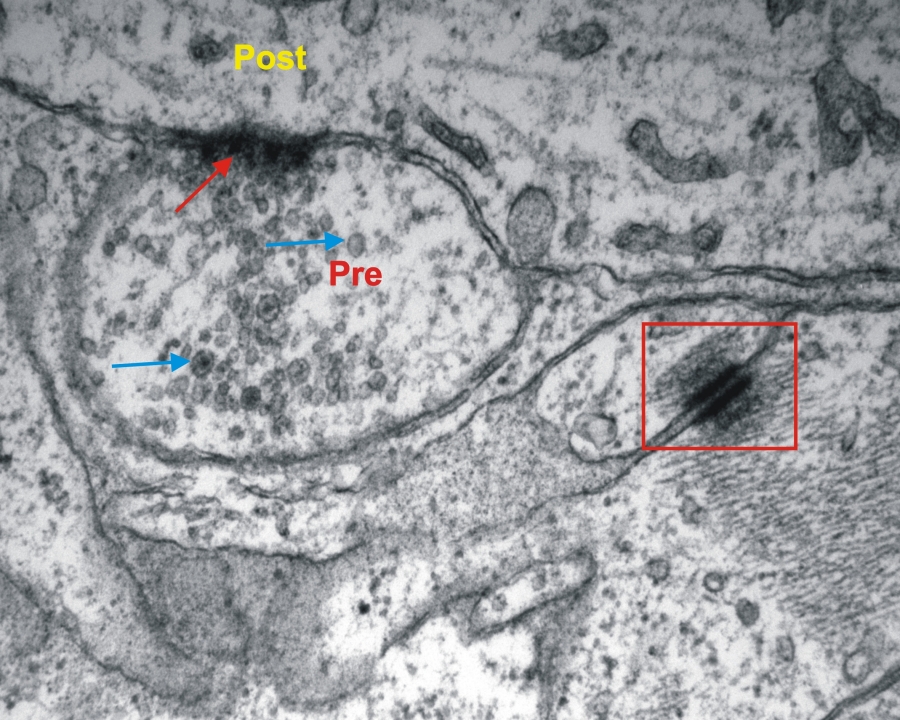
Transzmissziós elektronmikoszkópos felvétel entericus ganglionról patkány vékonybélben. (piros nyíl-kémiai szinapszis, Pre-preszinaptikus sejt, Post-posztszinaptikus sejt, kék nyíl-szinaptikus vezikulák, keret-sejtkapcsoló struktúra) 34000x
anterográd transzport
asztrocita
axo-axonikus szinapszis
axo-dendritikus szinapszis
axon
axo-szomatikus szinapszis
belső mezaxon
bipoláris neuron
chromogranin
dendrit
dynein
elektromos szinapszis
elsődleges érző neuron
ependimasejt
galaktocerebrozid
gap junction
gliasejt
Hortega-féle mikroglia
idegpálya
idegsejt
kémiai szinapszis
kinezin
külső mesaxon
lipofuszcin
lizoszóma
makroglia
mielinizáció
mikroglia
motorneuron
motoros véglemez
multipoláris neuron
nem fenesztrált endothelium
neuropilema
neurotranszmisszió
neurotranszmitter
Nissl-anyag
oligodendrocita
perifériás ideg
perikaryon
piramissejt
pituicita
posztszinaptikus sejt
preszinaptikus felszín
pszeudounipoláris neuron
Purkinje-sejt
Ranvier-féle befűződés
retrográd transzport
reziduális test
Schwann-sejt
szinaptofizin
szaltatorikus vezetés
szatellita sejt
szinapszis
szinaptikus rés
unipoláris neuron
vér-agy gát
vér-ganglion gát