Tartalom
A kötőszövetek sejtekből és a sejtek által termelt extracelluláris mátrixból épülnek fel. Az extracelluláris mátrixot kötőszöveti rostok, szerkezet nélküli alapállomány és szöveti nedvek alkotják. A kötőszöveti sejtek a koponya vázelemeinek jelentős részét kialakító dúcléc eredetű ektomezenchima sejtek kivételével a középső csíralemez mezenchima sejtjeiből differenciálódnak. Mivel egy adott kötőszövet funkcióját alapvetően meghatározza az, hogy benne milyen sejtek, milyen számban találhatók, valamint az, hogy ezek a sejtek milyen típusú extracelluláris mátrix komponenseket termelnek, a kötőszövetek morfológiai és funkcionális osztályozása is e paraméterek alapján történik.
A kötőszövetek osztályozása
Általános kötőszövetek
laza rostos kötőszövet
tömött rostos kötőszövet
rendezett
rendezetlen
Specializált kötőszövetek
zsírszövet
vérszövet
vérképző szövetek
nyirokszövet
Embrionális kötőszövetek
mezenchima
érett, kocsonyás kötőszövet
Kötőszöveti sejtek
A kötőszövetben található sejtek száma és mennyisége nagyon változó. Azok a sejtek, amelyek egy-egy kötőszövettípusban megjelennek, jól tükrözik az adott kötőszövet funkcióját. A sejtek számából viszont akár az egész szervezet funkcionális állapotára lehet következtetni. Jól ismert például az, hogy a laza rostos kötőszövet strukturális szerepén túl nagyon fontos szerepet játszik a szervezet immunológiai védelmében. Így egy gyulladásos folyamatban jelentősen megemelkedik benne azoknak a sejteknek a száma, amelyek a helyi véredényekből kivándorolva meghatározzák a szervezet immunológiai válaszreakcióit. A kötőszöveti sejteket két fő csoportra szokták osztani.
Az állandó sejtek közé tartoznak a fibroblastok, myofibroblastok, makrofágok, zsírsejtek, hízósejtek, differenciálatlan mesenchimasejtek.
A mobilis vagy vándorló kötőszöveti sejtek közé a limfociták, plazmasejtek, neutrofil, bazofil és eosinofil granulociták és monociták.
Az extracelluláris mátrix termeléséért elsősorban a fibroblastok felelősek. Rutin hematoxili-eosin festés után fibroblastokban csak a sejtmag látszik. Elektronmikroszkópos képeken nagyon sok fejlett DER és Golgi apparátus látható a citoplazmában, ami a sejtek magas szintetikus aktivitására utal.
A miofibroblasztok fibroblast és simaizomsejt sajátosságokkal is rendelkeznek. Elektronmikroszkópos képeken minden, a fibroblastokra is jellemző sejtorganellum megtalálható ezekben a sejtekben. Így kiterjedt DER, és sok Golgi látható bennük. Emellett azonban aktin filamentum kötegek és denz testek is gyakran vannak ezekben a sejtekben. A simaizomsejtekre jellemző bazális lamina azonban itt nem látható.
A szöveti makrofágok a vérben keringő monocitákból származnak. Amikor a monocita kikerül a keringési rendszerből, egy, az adott szövetre jellemző érési folyamaton megy keresztül és szöveti makrofággá alakul. Elektronmikroszkópos képeken a makrofágokon ujjszerű kitüremkedések láthatók, ezek a fagocitózis morfológiai jelei. Emellett egyéb struktúrák, mint pl. az endocitótikus vakuolák, lizoszómák szintén a fagocitózisra utalnak. A fejlett DER és Golgi rendszer jelenléte viszont a szintetikus tevékenységre utal.
A hízósejtek általában ovális sejtek, nagy kerek maggal. A citoplazmájuk nagy granulumokkal van tele, a sejtfelszínen mikrovillusok és membrángyűrődések láthatók. A citoplazmatikus sejtorganellumok száma nagyon alacsony. A hízósejtek rutin hisztológiai festékekkel nem igen láthatóak, viszont sok immunoreaktív anyag található bennük, így specifikus antitestek felhasználásával immunhisztokémiával jól vizsgálhatók.
A zsírsejtek fibroblastokból és differenciálatlan mezenchimasejtekből differenciálódnak, s citoplazmájukban fokozatosan zsírt halmoznak fel. Ha nagyon nagy számban jelennek meg, akkor kialakul a zsírszövet.
A differenciálatlan mesenchimasejtekből különböző kötőszöveti sejtek alakulhatnak ki. Ilyen differenciálatlan mesenchimasejteknek tekintjük pl. az erek körül elhelyezkedő pericitákat, amelyek pl. képesek simaizomsejtekké differenciálódni.
A limfociták elsősorban az immunválasz kialakításában játszanak szerepet. Kisebb számban minden laza rostos kötőszövetben megtalálhatók, de számuk gyulladásos folyamatok során megemelkedik. A limfocitáknak két alaptípusa van: a T limfociták, amelyek a celluláris-, valamint a B limfociták, amelyek a humorális immunválasz kialakításáért felelősek.
A plazmasejtek szintén állandó komponensei a laza rostos kötőszöveteknek, de gyulladásos folyamatok során ezek száma is megnő. Viszonylag nagy ovális sejtek, kiterjedt citoplazmával. A sejt erős bazofíliája utal a nagymennyiségű DER jelenlétére, amit az elektronmikroszkópos képek is igazolnak. A jelentős fehérjeszintetikus aktivitás ellenére a sejtmag erősen heterokromatikus. Ez valószínűleg azzal magyarázható, hogy ezek a sejtek általában egy, nagyon specifikus fehérjét (antitestet) termelnek, így a genomnak csak egy pici része aktív a szintézis folyamatában, vagyis a kromatin állomány nagyobbik része feltekeredett állapotban marad, aminek elektronmikroszkóposan is látható jele a heterokromatikus mag.
Erős gyulladásos reakciókban nagyon sok eozinofil és neutrofil granulocita vándorol a laza rostos kötőszövetbe. Más immunreakciókban a bazofil granulociták jelennek meg nagy tömegben. Ezek hisztamint szabadítanak fel, amelyeknek az allergiás bőrreakciók kialakulásában van fontos szerepe.
A sejtközötti állomány vagy extracelluláris mátrix
Az extracelluláris mátrix fő komponense a különböző kötőszöveti rostokból felépülő bonyolult hálózat, a főleg glükózaminoglikánokból (GAG) álló erősen hidrált gélszerű alapállomány, valamint különböző adhéziós molekulák, amelyek mint kapcsolómolekulák a kötőszövet elemeit bonyolult, összefüggő rendszerré szervezik.
A glükozaminoglikánok hosszú, el nem ágazó poliszaharid láncok, amelyek ismétlődő diszaharid egységekből épülnek fel. Egy egy egységet 70-200 diszaharid épít fel. Legfontosabb tulajdonságuk, hogy sok negatív töltést tartalmaznak, ezért a pozitív ionokat, mint pl. a Na+-ionokat vízburkukkal együtt megkötik, s így biztosítják a szövetek turgorját.
A hialuronsav kivételével mindegyik GAG képes kovalens kötésekkel fehérjékhez kapcsolódni, így proteoglikánok jönnek létre. Ezek mint óriásmolekulák, biztosítják a nagy hidrált terek kialakulását a kötőszövetek extracelluláris tereiben. A proteoglikánok térbeli elrendeződése és töltésük, valamint a nagy, összefüggő vízterek elősegítik a különböző anyagok szelektív diffúzióját. A tápanyagok és az oxigén az erekből a sejtek felé, míg az anyagcserevégtermékek a sejtekből az erek felé diffundálva biztosítják a szervezet működését. A GAG molekulákat szerkezetük szerint négy csoportra osztjuk: hialuronsav, chondroitin-szulfát és dermatán-szulfát, heparán-szulfát és heparin, keratán-szulfát. Ezek a különböző típusú kötőszövetekben különböző mennyiségban találhatók.
Rostos proteinek közül a kollagén, a fibrillin, az elasztin, és a fibronektin, amelyek meghatározó szerepet játszanak a különböző kötőszöveti rostok, s így az extracelluláris matrix struktúrájának és funkciójának a kialakításában.
A kollagén rostok mechanikai behatásokkal szemben rendkívül ellenállók. Gyakran alkotnak vékonyabb, vastagabb kötegeket, a fénymikroszkópos feloldás határán lévő fibrillumtól egészen a szabad szemmel is látható kollagénrostig. Rutin szövettani preparátumokon eozinnal rózsaszínűre festődnek. Főzés hatására az inakból, szalagokból kioldódnak és kocsonyás enyvet képeznek. Erről a sajátságról kapták a nevüket is, kolla-gen vagyis enyvadó rostok. A fénymikroszkóppal egyetlen rostnak látszó kollagénrost elektronmikroszkópos szinten elemi fibrillumokból álló köteg. Így a kollagénrost vastagsága a kötegben résztvevő elemi fibrillumok számától függ. Az elemi fibrillumok különböző vastagságú sötét és világos csíkokból álló jellegzetes harántcsíkolatot mutat. A csíkok mintázata 67 nanométerenként periódikusan ismétlődik.
A retikuláris vagy rácsrostok gyakran elágazódva, majd újra egyesülve finom térbeli hálózatot hoznak létre. Bár szinte minden laza rostos kötőszövetben előfordulnak, a legjellemzőbb előfordulási helyük az úgynevezett sejtképző szervekben, mint a lépben, a nyirokszervekben, csontvelőben van. Hematoxilin-eozin festéssel a rácsrostok nem különíthetők el a kollagénrostoktól. Az ezüst-sókkal azonban jól festhetők, ezért a rácsrostokat argirofil rostoknak is szokták nevezni.
A rugalmas vagy elasztikus rostok a kollagén rostoknál jóval vékonyabb, közel egyenletes vastagságú rostok. Rutin festési eljárásokkal nem festhetők, speciális, resorcin-fuchsin, vagy orcein festéssel tehetők láthatóvá. Rugalmas rostok a kötőszövetekben mindenhol előfordulnak, különösen ott, ahol a szervek működés közben erős alakváltozáson mennek keresztül. Elektronmikroszkópos képen a rostok átmetszetében egy elektrondenz központi mag látható, ez az elasztin, amihez 8-12 nm átmérőjű fibrillin mikrofibrillumok kapcsolódnak.
Az adhéziós molekulák a kötőszövet egyes elemeinek az összekapcsolását végzik. Az embrionális fejlődés korai szakaszában megjelennek, s már a gasztrulációs sejtmozgás során meghatározó szerepük van a sejtvándorlás irányításában, a csíralemezek kialakításában, az organogenezisben. A felnőtt szervezetben az adhéziós molekulák biztosítják azt, hogy a sejtek és a sejtközötti állomány különböző komponensei között a szövetek és az egész szervezet funkcionális állapotának megfelelő dinamikus egyensúlyi állapot fennmaradjon. A fibronektin, laminin és a tenaszcin fontos szerepet játszik a kötőszöveti sejtek és az extracelluláris matrix kapcsolódásában. Más adhéziós molekulák, mint pl. entaktin vagy trombospondin az extracelluláris matrix egyes elemeit képesek összekapcsolni, s így rendszerré alakítani azt.
A laza rostos kötőszövet erősen vaszkularizált szövet. A test külső és belső felszíneit burkoló hámréteg alatt helyezkedik el, s fontos szerepe van a véredényekkel nem rendelkező hámszövetek tápanyagokkal és oxigénnel történő ellátásában. Laza rostos kötőszövet található ezen kívül a mirigyparenchima alatt, illetve a kapillárisok körül is. A laza rostos kötőszövetben sok és sokféle sejt fordul elő. Specifikus stimulusok hatására a sejtek nagy része a helyi véredényekből vándorol ki, így ezek a sejtek csak átmenetileg találhatók meg a laza rostos kötőszövetben. Az extracelluláris mátrix komponensek közül itt a rostok jóval kisebb mennyiségben fordulnak elő, mint az alapállomány. A gélszerű alapállomány lehetővé teszi azt, hogy az erekből a tápanyagok és az oxigén a sejtekhez diffundáljon, s segíti azt is, hogy a sejtekben termelt különböző anyagcsere végtermékek könnyen visszajussanak az érrendszerbe.
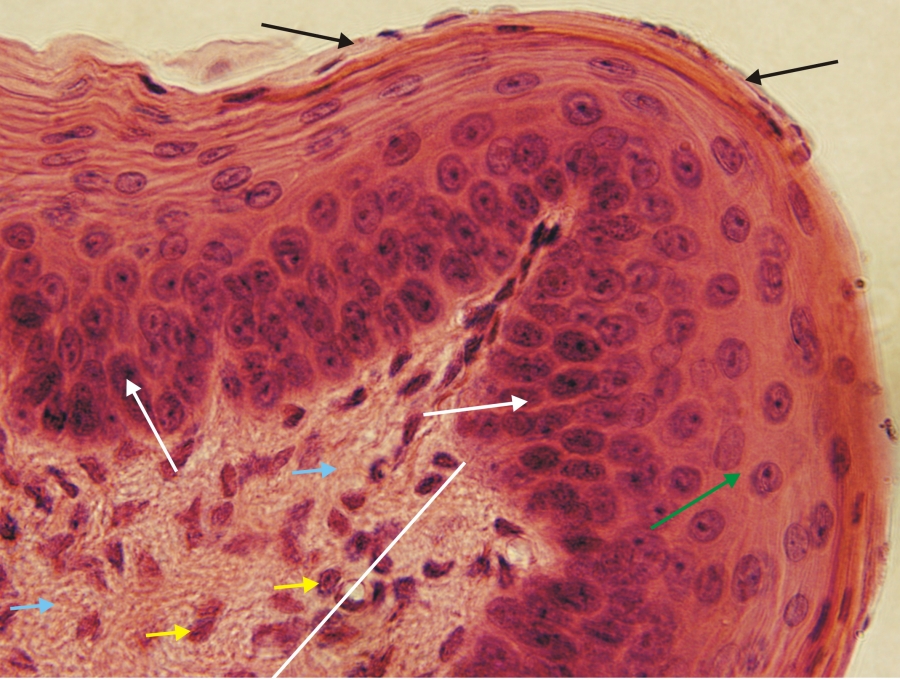
Fénymikroszkópos felvétel macska nyelvéből készült metszetről. (stratum basale-fehér nyíl, stratum polygonale-zöld nyíl, stratum planocellulare-fekete nyíl, fehér vonal-lazarostos kötőszövet, sárga nyilak-sejtmagok, kék nyilak-sejtközötti állomány). 1000x
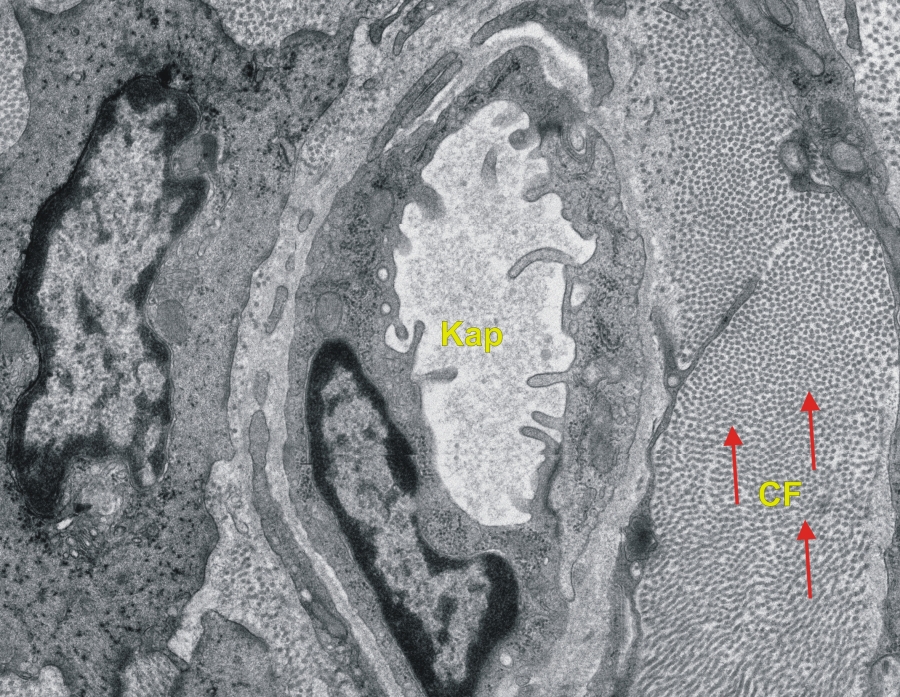
Transzmissziós elektronmikroszkópos felvétel patkány vékonybél kötőszövetéről. (CF-kollagénrostok, piros nyíl-kollagén rost keresztmetszete, Kap-kapilláris) 25000x
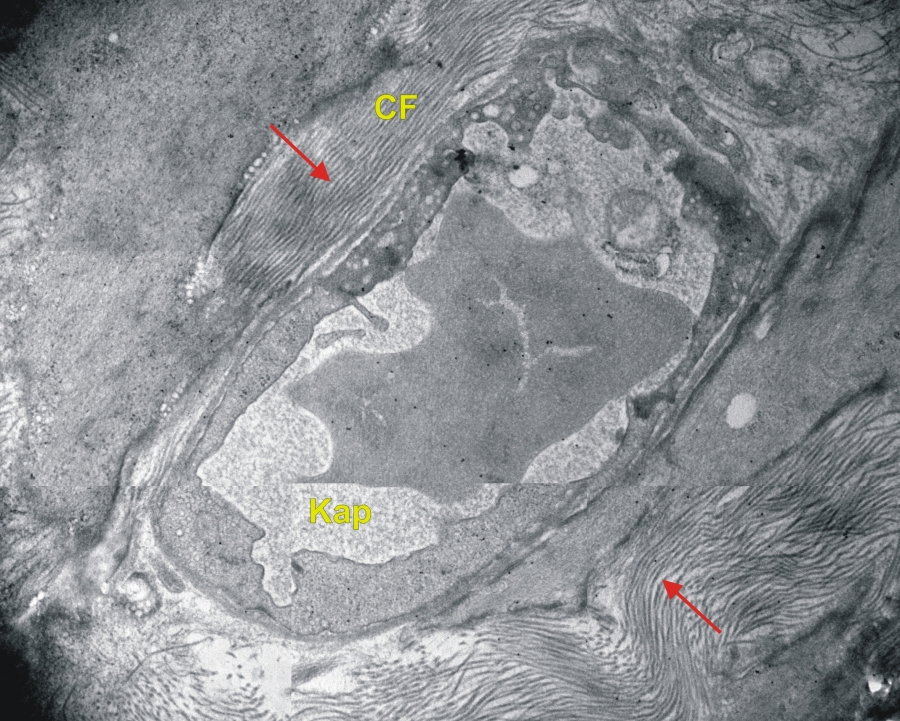
Transzmissziós elektronmikroszkópos felvétel patkány vékonybél kötőszövetéről. (CF-kollagén rostok, piros nyíl-kollagén rost hosszmetszet, Kap-kapilláris) 25000x
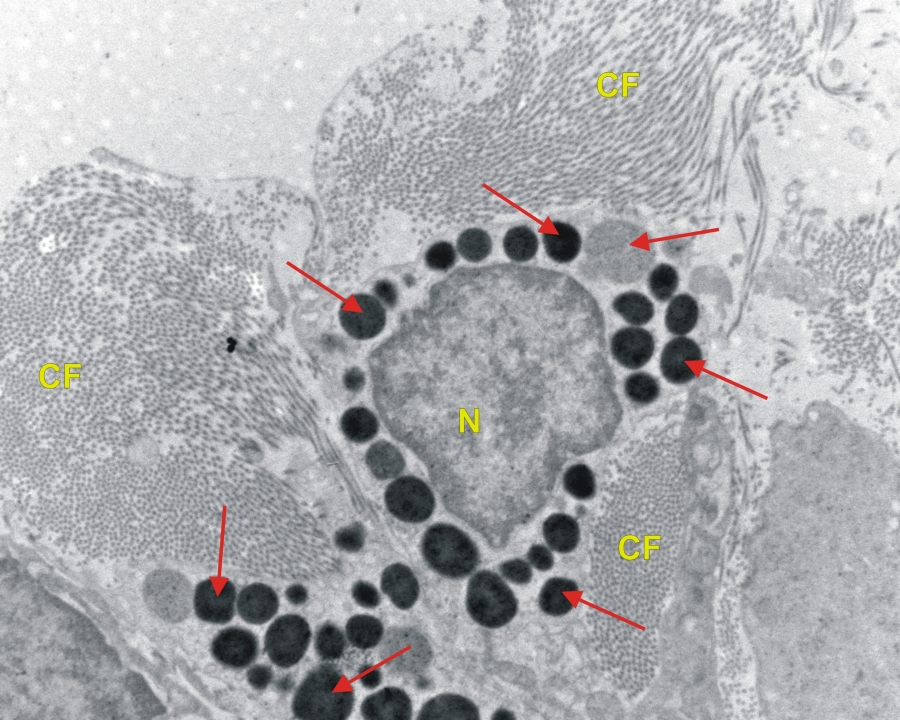
Transzmissziós elektronmikroszkópos felvlétel hízósejtekről patkány vékonybél kötőszövetes rétegében. (piros nyíl-citoplazmatikus granulum, N-sejtmag, CF-kollagén rostok) 5800x